


概要
薬学部ゲノム病態解析講座の寺尾知可史特任教授(理化学研究所生命医科学研究センター ゲノム解析応用研究チーム チームリーダー、静岡県立総合病院 臨床研究部免疫研究部長)、糖尿病?代謝ゲノム疾患研究チームの堀越桃子チームリーダーらの国際共同研究グループは、40歳から60歳の間に自然閉経した約20万人のヨーロッパ人女性対象にゲノムワイド関連解析(GWAS)[1]を行い、卵巣の加齢性変化に関わる290の疾患感受性領域(遺伝子座)[2]を同定しました。
本研究成果は、女性の生殖機能の維持や妊孕(よう)性[3]温存に対しての治療標的となり、女性の生殖可能期間延長のための治療法開発につながるものと期待できます。
今回、国際共同研究グループは、20万1323人のヨーロッパ人女性を対象にGWASを行い、自然閉経年齢(卵巣の加齢性変化)に関わる290の遺伝子座を同定しました。その結果から、早発卵巣不全(POI)[4]のリスクを予測できることを明らかにしました。さらに、それらの遺伝子座が幅広いDNA損傷応答(DDR)[5]プロセスにより生殖可能期間に関連していることが分かりました。
本研究は、科学雑誌『Nature』オンライン版(8月4日付)に掲載されました。
本研究成果は、女性の生殖機能の維持や妊孕(よう)性[3]温存に対しての治療標的となり、女性の生殖可能期間延長のための治療法開発につながるものと期待できます。
今回、国際共同研究グループは、20万1323人のヨーロッパ人女性を対象にGWASを行い、自然閉経年齢(卵巣の加齢性変化)に関わる290の遺伝子座を同定しました。その結果から、早発卵巣不全(POI)[4]のリスクを予測できることを明らかにしました。さらに、それらの遺伝子座が幅広いDNA損傷応答(DDR)[5]プロセスにより生殖可能期間に関連していることが分かりました。
本研究は、科学雑誌『Nature』オンライン版(8月4日付)に掲載されました。
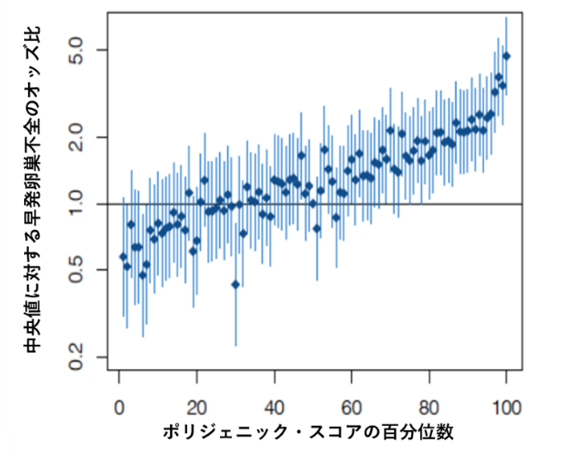
ポリジェニック?スコアと早発卵巣不全の関係
※国際共同研究グループ
理化学研究所 生命医科学研究センター
ゲノム解析応用研究チーム
チームリーダー 寺尾 知可史(てらお ちかし)
(静岡県立総合病院免疫 研究部長、欧洲杯押注平台_2024欧洲杯网站-官网app 特任教授)
糖尿病?代謝ゲノム疾患研究チーム
チームリーダー 堀越 桃子(ほりこし ももこ)
ケンブリッジ大学 MRC Epidemiologic Unit
ジョン?ペリー(John RB Perry)
研究支援
本研究は、日本医療研究開発機構(AMED)のオーダーメイド医療の実現プログラム「疾患関連遺伝子等の探索を効率化するための遺伝子多型情報の高度化(研究開発代表者:久保充明(当時))」を受けて行われました。本研究で使用したサンプルは、「オーダーメイド医療の実現プログラム」において収集されたものです。
理化学研究所 生命医科学研究センター
ゲノム解析応用研究チーム
チームリーダー 寺尾 知可史(てらお ちかし)
(静岡県立総合病院免疫 研究部長、欧洲杯押注平台_2024欧洲杯网站-官网app 特任教授)
糖尿病?代謝ゲノム疾患研究チーム
チームリーダー 堀越 桃子(ほりこし ももこ)
ケンブリッジ大学 MRC Epidemiologic Unit
ジョン?ペリー(John RB Perry)
研究支援
本研究は、日本医療研究開発機構(AMED)のオーダーメイド医療の実現プログラム「疾患関連遺伝子等の探索を効率化するための遺伝子多型情報の高度化(研究開発代表者:久保充明(当時))」を受けて行われました。本研究で使用したサンプルは、「オーダーメイド医療の実現プログラム」において収集されたものです。
発表内容
1. 背景
過去150年の間に日本女性の平均寿命は45歳から85歳に延びましたが注1)、閉経年齢は50~52歳で変化していません注2)。卵子の持つ遺伝子の健全性は年齢とともに減少し、自然な生殖能力は閉経の約10年前(つまり40歳?42歳)に停止します。近年は高齢出産を選択する女性が増えており、体外受精などの不妊治療や、卵子のもととなる卵母細胞や卵巣組織の凍結保存を行う女性が増えています注3,4)。
ところが、卵子や卵巣組織の採取は侵襲性が高い上に、凍結された成熟卵子を融解して体外受精に用いる場合、妊娠する確率は6.5%程度であり、母体年齢が高いほど妊娠の確率は低下するという問題があります注5)。
しかし、その生物学的メカニズムや、生殖能力を長く維持するための治療法についてはよく分かっていません。
注1)Christensen, K., Doblhammer, G., Rau, R. & Vaupel, J. W. Ageing populations: the challenges ahead. Lancet (London, England) 2009; 374, 1196–208.
注2)InterLACE Study Team. Variations in reproductive events across life: a pooled analysis of data from 505 147 women across 10 countries. Hum. Reprod. 2019; 34, 881–893.
注3)Donnez, J. & Dolmans, M.-M. Fertility Preservation in Women. N. Engl. J. Med. 2017; 377, 1657–1665.
注4)Yding Andersen, C., Mamsen, L. S. & Kristensen, S. G. FERTILITY PRESERVATION: Freezing of ovarian tissue and clinical opportunities. Reproduction 2019; 158, F27–F34.
注5)Argyle, C. E., Harper, J. C. & Davies, M. C. Oocyte cryopreservation: where are we now? Hum. Reprod. Update 2016; 22, 440–9.
ところが、卵子や卵巣組織の採取は侵襲性が高い上に、凍結された成熟卵子を融解して体外受精に用いる場合、妊娠する確率は6.5%程度であり、母体年齢が高いほど妊娠の確率は低下するという問題があります注5)。
しかし、その生物学的メカニズムや、生殖能力を長く維持するための治療法についてはよく分かっていません。
注1)Christensen, K., Doblhammer, G., Rau, R. & Vaupel, J. W. Ageing populations: the challenges ahead. Lancet (London, England) 2009; 374, 1196–208.
注2)InterLACE Study Team. Variations in reproductive events across life: a pooled analysis of data from 505 147 women across 10 countries. Hum. Reprod. 2019; 34, 881–893.
注3)Donnez, J. & Dolmans, M.-M. Fertility Preservation in Women. N. Engl. J. Med. 2017; 377, 1657–1665.
注4)Yding Andersen, C., Mamsen, L. S. & Kristensen, S. G. FERTILITY PRESERVATION: Freezing of ovarian tissue and clinical opportunities. Reproduction 2019; 158, F27–F34.
注5)Argyle, C. E., Harper, J. C. & Davies, M. C. Oocyte cryopreservation: where are we now? Hum. Reprod. Update 2016; 22, 440–9.
2. 研究手法と成果
国際共同研究グループはまず、40歳から60歳の間に自然閉経した20万1323人のヨーロッパ人女性のゲノムを用いてゲノムワイド関連解析(GWAS)を行いました。この結果、自然閉経年齢(卵巣の加齢性変化)と関連する290領域の遺伝子座を同定しました。さらに、バイオバンク?ジャパン[6]に登録されている日本人女性4万7140人を含む閉経年齢のデータから、一塩基多型(SNP)[7]と閉経年齢の関連の強さについて再現性を確認したところ、290領域のうち多くが再現されていましたが、いくつかの領域では関連の強さを示す効果量とアレル頻度[8]に人種による大きな違いがありました。
また、SNPの効果量を足し合わせて計算するポリジェニック?スコア(PGS)[9]を用いて、40歳未満で生殖機能低下が現れる早発卵巣不全(POI)を予測できるか調べました。すると、PGSの上位1%は、第50百分位数(中央値)に対するオッズ比(発症リスクの指標)が4.71を示し、POIの予測が可能であることが分かりました(図1)。これはPOIの原因遺伝子の一つであるFMR1遺伝子[10]に変異を持つ女性のリスクと同等であることも分かりました。
また、SNPの効果量を足し合わせて計算するポリジェニック?スコア(PGS)[9]を用いて、40歳未満で生殖機能低下が現れる早発卵巣不全(POI)を予測できるか調べました。すると、PGSの上位1%は、第50百分位数(中央値)に対するオッズ比(発症リスクの指標)が4.71を示し、POIの予測が可能であることが分かりました(図1)。これはPOIの原因遺伝子の一つであるFMR1遺伝子[10]に変異を持つ女性のリスクと同等であることも分かりました。
図1 ポリジェニック?スコアと早発卵巣不全の関係
ポリジェニック?スコア百分位数とその中央値に対する早発卵巣不全のオッズ比をプロット。上位1%の中央値に対するオッズ比は4.71であった。
今回ヒトで同定された遺伝子座は、幅広いDNA損傷応答(DDR)プロセスに関与しており、主要なDDR遺伝子の機能喪失型変異も含まれていました。マウスモデルでの実験から、これらのDDRプロセスが生涯にわたって作用し、卵巣予備能(卵巣に残っている卵子数)とその機能喪失率に関係することが明らかになりました。今回のGWASで検出された遺伝子のうちCHEK2遺伝子は減数分裂で修復されなかったDNA二本鎖切断や誘導されたDNA二本鎖切断を持つマウスの卵母細胞を淘汰するのに重要な役割を果たすことが知られています注6)。CHEK2遺伝子の機能をより明らかにするため、Chek2遺伝子をノックアウトしたマウスを観察したところ、卵巣予備能が長く維持され生殖可能期間が延長することが分かりました。
また、同定されたSNPを用いてメンデルランダム化[11]という因果関係を推定する解析により、女性の生殖可能期間の延長は骨の健康状態を改善し、2型糖尿病[12]のリスクを低下させる一方で、ホルモン感受性のある乳がん、卵巣がん、子宮内膜がんのリスクを高めることが示されました。
注6) Bolcun-Filas, E., Rinaldi, V. D., White, M. E. & Schimenti, J. C. Reversal of female infertility by Chk2 ablation reveals the oocyte DNA damage checkpoint pathway. Science 2014; 343, 533–536.
今回ヒトで同定された遺伝子座は、幅広いDNA損傷応答(DDR)プロセスに関与しており、主要なDDR遺伝子の機能喪失型変異も含まれていました。マウスモデルでの実験から、これらのDDRプロセスが生涯にわたって作用し、卵巣予備能(卵巣に残っている卵子数)とその機能喪失率に関係することが明らかになりました。今回のGWASで検出された遺伝子のうちCHEK2遺伝子は減数分裂で修復されなかったDNA二本鎖切断や誘導されたDNA二本鎖切断を持つマウスの卵母細胞を淘汰するのに重要な役割を果たすことが知られています注6)。CHEK2遺伝子の機能をより明らかにするため、Chek2遺伝子をノックアウトしたマウスを観察したところ、卵巣予備能が長く維持され生殖可能期間が延長することが分かりました。
また、同定されたSNPを用いてメンデルランダム化[11]という因果関係を推定する解析により、女性の生殖可能期間の延長は骨の健康状態を改善し、2型糖尿病[12]のリスクを低下させる一方で、ホルモン感受性のある乳がん、卵巣がん、子宮内膜がんのリスクを高めることが示されました。
注6) Bolcun-Filas, E., Rinaldi, V. D., White, M. E. & Schimenti, J. C. Reversal of female infertility by Chk2 ablation reveals the oocyte DNA damage checkpoint pathway. Science 2014; 343, 533–536.
3. 今後の期待
今回の解析で同定されたSNPからPGSを計算することで、POIを予測できることが示されました。さらに生殖可能期間と健康指標の間に因果関係があることが推測されました。また、解析で同定された遺伝子やパスウェイ[13]からDDRプロセスが生殖可能期間に関連していることが示されました。
今後、そのメカニズムをさらに解明することで、女性の生殖機能の維持や妊孕(よう)性温存に対しての治療標的となることが期待できます。
今後、そのメカニズムをさらに解明することで、女性の生殖機能の維持や妊孕(よう)性温存に対しての治療標的となることが期待できます。
4. 論文情報
<タイトル>
Genetic insights into the biological mechanisms governing human ovarian ageing
<著者名>
Katherine S Ruth, Felix R Day, Jazib Hussain, Ana Martínez-Marchal, Catherine E Aiken, Ajuna Azad, Deborah J Thompson, Lucie Knoblochovaz, Hironori Abe, Jane L Tarry-Adkins, Javier Martin Gonzalez, Pierre Fontanillas, Annique Claringbould, Olivier B Bakker, Patrick Sulem, Robin G Walters, Chikashi Terao, Sandra Turon, Momoko Horikoshi, Kuang Lin, N Charlotte Onland-Moret, Emil Peter Trane Hertz, Pascal N Timshel, Vallari Shukla, Rehannah Borup, Kristina W Olsen, Paula Aguilera, Mònica Ferrer-Roda, Yan Huang, Stasa Stankovic, Paul RHJ Timmers, Thomas U Ahearn, Behrooz Z Alizadeh, Elnaz Naderi, Irene L Andrulis, Alice M Arnold, Kristan J Aronson, Annelie Augustinsson, Stefania Bandinelli, Caterina M Barbieri, Robin N Beaumont, Heiko Becher, Matthias W Beckmann, Stefania Benonisdottir, Sven Bergmann, Murielle Bochud, Eric Boerwinkle, Stig E Bojesen, Manjeet K Bolla, Dorret I Boomsma, Nicholas Bowker, Jennifer A Brody, Linda Broer, Julie E Buring, Archie Campbell, Harry Campbell, Jose E Castelao, Eulalia Catamo, Stephen J Chanock, Georgia Chenevix-Trench, Marina Ciullo Tanguy Corre, Fergus J Couch, Angela Cox, Laura Crisponi, Simon S Cross, Francesco Cucca, Kamila Czene, George Davey-Smith, Eco JCN de Geus, Renée de Mutsert, Immaculata De Vivo, Ellen W Demerath, Joe Dennis, Alison M Dunning, Miriam Dwek, Mikael Eriksson, T?nu Esko, Peter A Fasching, Jessica D Faul, Luigi Ferrucci, Nora Franceschini, Timothy M Frayling, Manuela Gago-Dominguez, Massimo Mezzavilla, Montserrat García-Closas, Christian Gieger, Graham G Giles, Harald Grallert, Daniel F Gudbjartsson, Vilmundur Gudnason, Pascal Guénel, Christopher A Haiman, Niclas H?kansson, Per Hall, Caroline Hayward, Chunyan He, Wei He, Gerardo Heiss, Miya K H?ffding, John L Hopper, Jouke J Hottenga, Frank Hu, David Hunter, Mohammad A Ikram, Rebecca D Jackson, Micaella DR Joaquim, Esther M John, Peter K Joshi, David Karasik, Sharon LR Kardia, Christiana Kartsonaki, Robert Karlsson, Cari M Kitahar, Ivana Kolcic, Charles Kooperberg, Peter Kraft, Allison W Kurian, Zoltan Kutalik, Martina La Bianca, Genevieve LaChance, Claudia Langenberg, Lenore J Launer, Joop SE Laven, Deborah A Lawlor, Loic Le Marchand, Jingmei Li, Annika Lindblom, Sara Lindstrom, Tricia Lindstrom, Martha Linet, YongMei Liu, Simin Liu, Jian'an Luan, Reedik M?gi, Patrik KE Magnusson, Massimo Mangino, Arto Mannermaa, Brumat Marco, Jonathan Marten, Nicholas G Martin, Hamdi Mbarek, Barbara McKnight, Sarah E Medland, Christa Meisinger, Thomas Meitinger, Cristina Menni, Andres Metspalu, Lili Milani, Roger L Milne, Grant W Montgomery, Dennis O Mook-Kanamori, Antonella Mulas, Anna M Mulligan, Alison Murray, Mike A Nalls, Anne Newman, Raymond Noordam, Teresa Nutile, Dale R Nyholt, Andrew F Olshan, H?kan Olsson, Jodie N Painter, Alpa V Patel, Nancy L Pedersen, Natalia Perjakova, Annette Peters, Ulrike Peters, Paul DP Pharoah, Ozren Polasek, Eleonora Porcu, Bruce M Psaty, Iffat Rahman, Gad Rennert, Hedy S Rennert, Paul M Ridker, Susan M Ring, Antonietta Robino, Lynda M Rose, Frits R Rosendaal, Jacques Rossouw, Igor Rudan, Rico Rueedi, Daniela Ruggiero, Cinzia F Sala, Emmanouil Saloustros, Dale P Sandler, Serena Sanna, Elinor J Sawyer, Chloé Sarnowski, David Schlessinger, Marjanka K Schmidt, Minouk J Schoemaker, Katharina E Schraut, Christopher Scott, Saleh Shekari1, Amruta Shrikhande3, Albert V Smith, Blair H Smith, Jennifer A Smith, Rossella Sorice, Melissa C Southey, Tim D Spector, John J Spinelli, Meir Stampfer, Doris St?ckl, Joyce BJ van Meurs, Konstantin Strauch, Unnur Styrkarsdottir, Anthony J Swerdlow, Toshiko Tanaka, Lauren R Teras, Alexander Teumer, Unnur ?orsteinsdottir, Nicholas J Timpson, Daniela Toniolo, Michela Traglia, Melissa A Troester, Thérèse Truong, Jessica Tyrrell1, André G Uitterlinden, Sheila Ulivi, Celine M Vachon, Veronique Vitart, Uwe V?lker, Peter Vollenweider, Henry V?lzke, Qin Wang, Nicholas J Wareham, Clarice R Weinberg, David R Weir, Amber N Wilcox, Ko Willems van Dijk, Gonneke Willemsen, James F Wilson, Bruce HR Wolffenbuttel, Alicja Wolk, Andrew R Wood1, Wei Zhao, Marek Zygmunt, Biobank-based Integrative Omics Study (BIOS) Consortium, eQTLGen Consortium, The Biobank Japan Project, China Kadoorie Biobank Collaborative Group, kConFab Investigators, The LifeLines Cohort Study, The InterAct consortium, 23andMe Research Team, Zhengming Chen, Liming Li, Lude Franke, Stephen Burgess, Patrick Deelen, Tune H Pers, Marie Louise Gr?ndahl, Claus Yding Andersen, Anna Pujol, Andres J Lopez-Contreras, Jeremy A Daniel, Kari Stefansson, Jenny Chang-Claude, Yvonne T van der Schouw, Kathyrn L Lunetta, Daniel I Chasman, Douglas F Easton, Jenny A Visser, Susan E Ozanne, Satoshi H Namekawa, Petr Solc, Joanne M Murabito, Ken K Ong, Eva R Hoffmann, Anna Murray, Ignasi Roig and John RB Perry
<雑誌>
Nature
<DOI>
10.1038/s41586-021-03779-7
Genetic insights into the biological mechanisms governing human ovarian ageing
<著者名>
Katherine S Ruth, Felix R Day, Jazib Hussain, Ana Martínez-Marchal, Catherine E Aiken, Ajuna Azad, Deborah J Thompson, Lucie Knoblochovaz, Hironori Abe, Jane L Tarry-Adkins, Javier Martin Gonzalez, Pierre Fontanillas, Annique Claringbould, Olivier B Bakker, Patrick Sulem, Robin G Walters, Chikashi Terao, Sandra Turon, Momoko Horikoshi, Kuang Lin, N Charlotte Onland-Moret, Emil Peter Trane Hertz, Pascal N Timshel, Vallari Shukla, Rehannah Borup, Kristina W Olsen, Paula Aguilera, Mònica Ferrer-Roda, Yan Huang, Stasa Stankovic, Paul RHJ Timmers, Thomas U Ahearn, Behrooz Z Alizadeh, Elnaz Naderi, Irene L Andrulis, Alice M Arnold, Kristan J Aronson, Annelie Augustinsson, Stefania Bandinelli, Caterina M Barbieri, Robin N Beaumont, Heiko Becher, Matthias W Beckmann, Stefania Benonisdottir, Sven Bergmann, Murielle Bochud, Eric Boerwinkle, Stig E Bojesen, Manjeet K Bolla, Dorret I Boomsma, Nicholas Bowker, Jennifer A Brody, Linda Broer, Julie E Buring, Archie Campbell, Harry Campbell, Jose E Castelao, Eulalia Catamo, Stephen J Chanock, Georgia Chenevix-Trench, Marina Ciullo Tanguy Corre, Fergus J Couch, Angela Cox, Laura Crisponi, Simon S Cross, Francesco Cucca, Kamila Czene, George Davey-Smith, Eco JCN de Geus, Renée de Mutsert, Immaculata De Vivo, Ellen W Demerath, Joe Dennis, Alison M Dunning, Miriam Dwek, Mikael Eriksson, T?nu Esko, Peter A Fasching, Jessica D Faul, Luigi Ferrucci, Nora Franceschini, Timothy M Frayling, Manuela Gago-Dominguez, Massimo Mezzavilla, Montserrat García-Closas, Christian Gieger, Graham G Giles, Harald Grallert, Daniel F Gudbjartsson, Vilmundur Gudnason, Pascal Guénel, Christopher A Haiman, Niclas H?kansson, Per Hall, Caroline Hayward, Chunyan He, Wei He, Gerardo Heiss, Miya K H?ffding, John L Hopper, Jouke J Hottenga, Frank Hu, David Hunter, Mohammad A Ikram, Rebecca D Jackson, Micaella DR Joaquim, Esther M John, Peter K Joshi, David Karasik, Sharon LR Kardia, Christiana Kartsonaki, Robert Karlsson, Cari M Kitahar, Ivana Kolcic, Charles Kooperberg, Peter Kraft, Allison W Kurian, Zoltan Kutalik, Martina La Bianca, Genevieve LaChance, Claudia Langenberg, Lenore J Launer, Joop SE Laven, Deborah A Lawlor, Loic Le Marchand, Jingmei Li, Annika Lindblom, Sara Lindstrom, Tricia Lindstrom, Martha Linet, YongMei Liu, Simin Liu, Jian'an Luan, Reedik M?gi, Patrik KE Magnusson, Massimo Mangino, Arto Mannermaa, Brumat Marco, Jonathan Marten, Nicholas G Martin, Hamdi Mbarek, Barbara McKnight, Sarah E Medland, Christa Meisinger, Thomas Meitinger, Cristina Menni, Andres Metspalu, Lili Milani, Roger L Milne, Grant W Montgomery, Dennis O Mook-Kanamori, Antonella Mulas, Anna M Mulligan, Alison Murray, Mike A Nalls, Anne Newman, Raymond Noordam, Teresa Nutile, Dale R Nyholt, Andrew F Olshan, H?kan Olsson, Jodie N Painter, Alpa V Patel, Nancy L Pedersen, Natalia Perjakova, Annette Peters, Ulrike Peters, Paul DP Pharoah, Ozren Polasek, Eleonora Porcu, Bruce M Psaty, Iffat Rahman, Gad Rennert, Hedy S Rennert, Paul M Ridker, Susan M Ring, Antonietta Robino, Lynda M Rose, Frits R Rosendaal, Jacques Rossouw, Igor Rudan, Rico Rueedi, Daniela Ruggiero, Cinzia F Sala, Emmanouil Saloustros, Dale P Sandler, Serena Sanna, Elinor J Sawyer, Chloé Sarnowski, David Schlessinger, Marjanka K Schmidt, Minouk J Schoemaker, Katharina E Schraut, Christopher Scott, Saleh Shekari1, Amruta Shrikhande3, Albert V Smith, Blair H Smith, Jennifer A Smith, Rossella Sorice, Melissa C Southey, Tim D Spector, John J Spinelli, Meir Stampfer, Doris St?ckl, Joyce BJ van Meurs, Konstantin Strauch, Unnur Styrkarsdottir, Anthony J Swerdlow, Toshiko Tanaka, Lauren R Teras, Alexander Teumer, Unnur ?orsteinsdottir, Nicholas J Timpson, Daniela Toniolo, Michela Traglia, Melissa A Troester, Thérèse Truong, Jessica Tyrrell1, André G Uitterlinden, Sheila Ulivi, Celine M Vachon, Veronique Vitart, Uwe V?lker, Peter Vollenweider, Henry V?lzke, Qin Wang, Nicholas J Wareham, Clarice R Weinberg, David R Weir, Amber N Wilcox, Ko Willems van Dijk, Gonneke Willemsen, James F Wilson, Bruce HR Wolffenbuttel, Alicja Wolk, Andrew R Wood1, Wei Zhao, Marek Zygmunt, Biobank-based Integrative Omics Study (BIOS) Consortium, eQTLGen Consortium, The Biobank Japan Project, China Kadoorie Biobank Collaborative Group, kConFab Investigators, The LifeLines Cohort Study, The InterAct consortium, 23andMe Research Team, Zhengming Chen, Liming Li, Lude Franke, Stephen Burgess, Patrick Deelen, Tune H Pers, Marie Louise Gr?ndahl, Claus Yding Andersen, Anna Pujol, Andres J Lopez-Contreras, Jeremy A Daniel, Kari Stefansson, Jenny Chang-Claude, Yvonne T van der Schouw, Kathyrn L Lunetta, Daniel I Chasman, Douglas F Easton, Jenny A Visser, Susan E Ozanne, Satoshi H Namekawa, Petr Solc, Joanne M Murabito, Ken K Ong, Eva R Hoffmann, Anna Murray, Ignasi Roig and John RB Perry
<雑誌>
Nature
<DOI>
10.1038/s41586-021-03779-7
5. 補足説明
[1] ゲノムワイド関連解析(GWAS)
形質に対する遺伝的関連を知るための手法であり、SNP(一塩基多型)を用いて解析するものが一般的である。形質(疾患のある/なしや量的形質)を目的変数、SNPの量的情報や各種共変量を説明変数にしてモデル化し、SNPの関連を評価する。GWASはGenome-Wide Association Studyの略。
[2] 疾患感受性領域(遺伝子座)
疾患の発症に関連している染色体上の領域のこと。
[3] 妊孕(よう)性
生殖機能とほぼ同義とされ、男女における妊娠に必要な臓器、配偶子、機能のこと。
[4] 早発卵巣不全(POI)
40歳より前に月経が3ヵ月以上来なくなり、卵巣機能が低下した状態のこと。早発卵巣不全になると排卵をしなくなるため妊娠が非常に困難になり、エストロゲンが不足?低下することによって骨折しやすくなるなど、全身の健康に影響が及ぶ。海外の報告では、女性100人あたり1人程度の割合で発症するというデータもある。POIはPremature Ovarian Insufficiencyの略。
[5] DNA損傷応答(DDR)
細胞は外因性および内因性ストレス因子にさらされており、これらのストレス因子によってDNA損傷が起こると、DNA損傷応答が誘導され、多くのDNA修復経路が活性化される。DNA損傷が適切に修復されない場合、突然変異が起こるなどしてゲノムの不安定性が広がっていく。DDRはDNA Damage Responseの略。
[6] バイオバンク?ジャパン(BBJ)
日本人集団27万人を対象とした生体試料のバイオバンクで、東京大学医科学研究所内に設置されている。理化学研究所が実験を行って取得した約20万人のゲノムデータを保有する。オーダーメイド医療の実現プログラムを通じて実施され、ゲノムDNAや血清サンプルを臨床情報とともに収集し、研究者へのデータ提供や分譲を行っている。
[7] 一塩基多型(SNP)
一つの遺伝的座位に、二つかそれ以上の頻度の高い異なるアレルが存在する状態のことを遺伝的多型という。一つの塩基がほかの塩基に変わる多型を、一塩基多型と呼ぶ。SNPはSingle Nucleotide Polymorphismの略。
[8] アレル頻度
個々のヒトゲノムを比較すると、染色体上の場所が同一であっても、遺伝子や個々の塩基配列が異なる場合がある。これらの遺伝子や塩基配列を、アレルという。例えば、ある染色体上の位置において、個人によりAA/AG/GGなどA(アデニン)やG(グアニン)の頻度をアレル頻度という。
[9] ポリジェニック?スコア(PGS)
生活習慣病やがんといった多因子疾患においては、GWASによって病気発症リスクと関連する多くのSNPが発見されている。しかし、これらのSNPの遺伝的効果は小さく(1.05~2倍)、多因子疾患は、遺伝的効果の少ない多数のSNPが蓄積して疾患リスクを高めていると考えられている。PGSはGWASによって発見された病気と関連するSNPを複数個、または測定した全てのSNPの情報をスコア化した指数のことで、個人の病気の発症リスクを推定する方法の一つである。PGSはPolygenic scoreの略。
[10] FMR1遺伝子
FMR1遺伝子はX染色体に存在し、第1エクソンの非翻訳領域にCGG反復配列がある。反復数が60-200の場合をpremutationと呼び、これを持つ女性の約20%で早発卵巣不全(PO))を発症する。単一遺伝子変異によるPOIの原因遺伝子の一つ。
[11] メンデルランダム化
暴露に関連するSNPを操作変数として用いて、暴露因子と疾病との間の因果関係について推測する解析手法である。これには遺伝子多型が暴露と関連していること、遺伝子多型が暴露を介してのみ疾病の発症に影響すること、遺伝子多型とがんの発生との間に交絡因子が存在しないこと、の三つの前提条件を満たす必要がある。
[12] 2型糖尿病
糖尿病は、大きく1型糖尿病、2型糖尿病、その他の特定の機序や疾患による糖尿病、妊娠糖尿病の四つに分類されている。糖尿病の大部分を占める2型糖尿病は、インスリン分泌低下とインスリン抵抗性(インスリンの働きが悪くなること)が合わさり、血糖値が上昇し、発症する。発症には、遺伝因子(家系)と環境因子(過食、肥満、運動不足などの生活習慣)の両者が深く関わっている。
[13] パスウェイ
遺伝子やタンパク質の相互作用を経路図として表したもの。
形質に対する遺伝的関連を知るための手法であり、SNP(一塩基多型)を用いて解析するものが一般的である。形質(疾患のある/なしや量的形質)を目的変数、SNPの量的情報や各種共変量を説明変数にしてモデル化し、SNPの関連を評価する。GWASはGenome-Wide Association Studyの略。
[2] 疾患感受性領域(遺伝子座)
疾患の発症に関連している染色体上の領域のこと。
[3] 妊孕(よう)性
生殖機能とほぼ同義とされ、男女における妊娠に必要な臓器、配偶子、機能のこと。
[4] 早発卵巣不全(POI)
40歳より前に月経が3ヵ月以上来なくなり、卵巣機能が低下した状態のこと。早発卵巣不全になると排卵をしなくなるため妊娠が非常に困難になり、エストロゲンが不足?低下することによって骨折しやすくなるなど、全身の健康に影響が及ぶ。海外の報告では、女性100人あたり1人程度の割合で発症するというデータもある。POIはPremature Ovarian Insufficiencyの略。
[5] DNA損傷応答(DDR)
細胞は外因性および内因性ストレス因子にさらされており、これらのストレス因子によってDNA損傷が起こると、DNA損傷応答が誘導され、多くのDNA修復経路が活性化される。DNA損傷が適切に修復されない場合、突然変異が起こるなどしてゲノムの不安定性が広がっていく。DDRはDNA Damage Responseの略。
[6] バイオバンク?ジャパン(BBJ)
日本人集団27万人を対象とした生体試料のバイオバンクで、東京大学医科学研究所内に設置されている。理化学研究所が実験を行って取得した約20万人のゲノムデータを保有する。オーダーメイド医療の実現プログラムを通じて実施され、ゲノムDNAや血清サンプルを臨床情報とともに収集し、研究者へのデータ提供や分譲を行っている。
[7] 一塩基多型(SNP)
一つの遺伝的座位に、二つかそれ以上の頻度の高い異なるアレルが存在する状態のことを遺伝的多型という。一つの塩基がほかの塩基に変わる多型を、一塩基多型と呼ぶ。SNPはSingle Nucleotide Polymorphismの略。
[8] アレル頻度
個々のヒトゲノムを比較すると、染色体上の場所が同一であっても、遺伝子や個々の塩基配列が異なる場合がある。これらの遺伝子や塩基配列を、アレルという。例えば、ある染色体上の位置において、個人によりAA/AG/GGなどA(アデニン)やG(グアニン)の頻度をアレル頻度という。
[9] ポリジェニック?スコア(PGS)
生活習慣病やがんといった多因子疾患においては、GWASによって病気発症リスクと関連する多くのSNPが発見されている。しかし、これらのSNPの遺伝的効果は小さく(1.05~2倍)、多因子疾患は、遺伝的効果の少ない多数のSNPが蓄積して疾患リスクを高めていると考えられている。PGSはGWASによって発見された病気と関連するSNPを複数個、または測定した全てのSNPの情報をスコア化した指数のことで、個人の病気の発症リスクを推定する方法の一つである。PGSはPolygenic scoreの略。
[10] FMR1遺伝子
FMR1遺伝子はX染色体に存在し、第1エクソンの非翻訳領域にCGG反復配列がある。反復数が60-200の場合をpremutationと呼び、これを持つ女性の約20%で早発卵巣不全(PO))を発症する。単一遺伝子変異によるPOIの原因遺伝子の一つ。
[11] メンデルランダム化
暴露に関連するSNPを操作変数として用いて、暴露因子と疾病との間の因果関係について推測する解析手法である。これには遺伝子多型が暴露と関連していること、遺伝子多型が暴露を介してのみ疾病の発症に影響すること、遺伝子多型とがんの発生との間に交絡因子が存在しないこと、の三つの前提条件を満たす必要がある。
[12] 2型糖尿病
糖尿病は、大きく1型糖尿病、2型糖尿病、その他の特定の機序や疾患による糖尿病、妊娠糖尿病の四つに分類されている。糖尿病の大部分を占める2型糖尿病は、インスリン分泌低下とインスリン抵抗性(インスリンの働きが悪くなること)が合わさり、血糖値が上昇し、発症する。発症には、遺伝因子(家系)と環境因子(過食、肥満、運動不足などの生活習慣)の両者が深く関わっている。
[13] パスウェイ
遺伝子やタンパク質の相互作用を経路図として表したもの。
お問い合わせ
薬学部 寺尾 知可史
E-mail:cterao@u-shizuoka-ken.ac.jp
E-mail:cterao@u-shizuoka-ken.ac.jp

